

Selenium toxicity
Selenium is an essential micronutrient of many living species, including humans, because the selenocysteine amino acid is necessary for the activity of a few important antioxidant enzymes. At high doses selenium compounds become highly toxic. However, in the past decade, it was reported that supranutritional levels of selenium can afford benefices in cancer prevention and therapy. It should be noted that the gap between toxic and prophylactic or therapeutic doses of selenium is narrow.
Numerous studies, in a variety of model systems have been undertaken to define the underlying mechanisms supporting the effects of selenium in cellular toxicity or prevention of cancer. It has become apparent that different selenocompounds exert variable beneficial or toxic effects, depending on chemical form, concentration and type of cells used in the assay.
In this context, we have focused our efforts on the import, storage and toxicity of various selenium metabolites in the yeast Saccharomyces cerevisiae. Because this organism lacks the pathway for the specific incorporation of selenocysteine into proteins, interferences with the function of selenium incorporated in the active site of proteins is precluded. Secondly, yeast is a good model to study selenium toxicity, because interconversion of selenium metabolites can be abolished by using gene mutants, allowing to pinpoint metabolic routes specific to each compound.
Recent results
Mechanisms of hydrogen selenide toxicity (Peyroche, G., Saveanu, C., Dauplais, M., Lazard., M, Beuneu, F., Decourty, L., Malabat, C., Jacquier, A., Blanquet, S. & Plateau, P., Sodium selenide toxicity is mediated by O2-dependent DNA breaks. PLoS One 7, e36343 (2012) [Abstract] [Full text])
Our initial work on selenium, published in 2007, established that toxicity of selenite ions for yeast was significantly enhanced by the presence in culture media of thiol compounds and that extracellular production of hydrogen selenide (H2Se) accounted for this hypertoxicity (Tarze et al., J. Biol. Chem. 282, 8759-8767 (2007)).
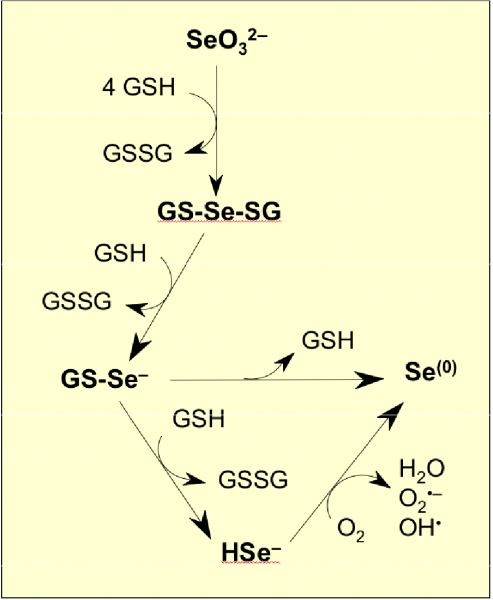
In vitro, glutathione spontaneously reacts with selenite (SeO32-) to form various selenocompounds: selenodiglutathione (GS-Se-SG), glutathioselenol (GS-Se-), hydrogen selenide (HSe-) and elemental selenium (Se(0))
This observation possibly explains why cancer cells resistant to conventional chemotherapy become remarkably sensitive to the presence of selenite. Indeed, these resistant cells often secrete increased amounts of cysteine. As a consequence, conversion of selenite to selenide (H2Se) would be promoted in the neighborhood of these cells and the toxicity of selenite would be enhanced (Olm, et al., Proc. Natl. Acad. Sci. USA 106, 11400-11405 (2009)).
In recent years, H2Se has emerged as a key intermediate in selenium metabolism. Therefore, we searched to decipher the causes of hydrogen selenide toxicity. For this purpose, we screened a library of yeast deletion mutants (in collaboration with the Institut Pasteur research unit of Genetics of Macromolecular Interactions) and showed that among the genes whose deletion caused hypersensitivity to H2Se, homologous recombination and DNA damage checkpoints genes were over-represented. This suggested that double-strand breaks in DNA were a major cause of selenide toxicity. Consistent with this hypothesis, treatment of Saccharomyces cerevisiae cells with hydrogen selenide triggered G2/M checkpoint activation and induced in vivo chromosome fragmentation, as shown by FACS and pulse-field gel electrophoresis experiments.
We also found that, in vitro, H2Se caused single-strand breaks in DNA, depending on the presence of O2. Hydrogen selenide is known to quickly and spontaneously react with dioxygen to form elemental selenium and water, and also reactive oxygen species (ROS). Inhibition of the formation of DNA breaks by selenide was observed in the presence of mannitol, a hydroxyl radical quencher, but no more when superoxide dismutase or catalase were added to the reaction buffer. In collaboration with the Irradiated Solids Laboratory (LSI) at the Ecole polytechnique, we detected by ESR experiments the characteristic •OH signature in an oxygenated solution of sodium selenide. Finally, we showed that, as could be expected, selenide toxicity strictly depended on the presence of dioxygen.
Altogether, these results demonstrate that, in yeast cells, hydrogen selenide induces toxic DNA breaks through an O2-dependent radical-based mechanism. Because of their increased frequency of division, tumor cells can be suspected to have less time than normal cells to repair such breaks. This behavior would account for a particular sensitivity of these cells to the presence of selenium.

Microarray DNA chip (detail) used to identify yeast genes whose deletion modifies the sensitivity to hydrogen selenide. Red: more sensitive. Green: less sensitive. Yellow: neutral
Neutralization by metal ions of the toxicity of sodium selenide (Dauplais, M., Lazard, M., Blanquet, S. & Plateau, P., Neutralization by metal ions of the toxicity of sodium selenide. PLoS One 8, e54353 (2013)[Abstract] [Full text])
Inert metal-selenide nanoparticles are found in animals. They are believed to result from the concomitant presence of the two chemicals and to afford cross-protection against the toxicities of both metals and selenocompounds. To systematically study the toxicities of metal salt and sodium selenide mixtures, we used the death rate of the yeast S. cerevisiae cells as an indicator. In parallel, we assessed the abilities of these mixtures to produce colloids. Studied metal cations could be classified in three groups: (i) metal ions that protect cells against selenium toxicity and form insoluble colloids with selenide (Ag+, Cd2+, Cu2+, Hg2+, Pb2+and Zn2+), (ii) metal ions which protect cells by producing insoluble metal-selenide complexes and/or by catalyzing hydrogen selenide oxidation in the presence of dioxygen (Co2+ and Ni2+) and, finally, (iii) metal ions which do not afford protection and do not interact (Ca2+, Mg2+, Mn2+) or weakly interact (Fe2+) with selenide.
This study highlights the potential importance of metals on the selenium status of individuals. As mentioned above, selenium might be a valuable food additive to prevent some cancers. However, results of trials comparing the cancer incidence rate among selenium-exposed and control individuals are much debated.Variations in selenium bioavailability from one individual to another may increase the complexity background of these trials. Clearly, consumption with dietary selenium of uncontrolled levels of metals and storage of metal-selenium nanoparticles can contribute to this difficulty.
Selenodigluthathione uptake by the Saccharomyces cerevisiae vacuolar transporter Ycf1p (Lazard, M., Ha-Duong, N. T., Mounié, S., Perrin, R., Plateau, P. & Blanquet, S., Selenodiglutathione uptake by theSaccharomyces cerevisiae vacuolar ATP-binding cassette transporter Ycf1p. FEBS J. 278, 4112-4121 (2011) [Abstract])
The Saccharomyces cerevisiae vacuolar ABC transporter Ycf1p is involved in heavy metal detoxification by mediating the ATP-dependent transport of glutathione-metal conjugates to the vacuole. A positive role of Ycf1p in selenium detoxification was therefore expected. However, Pinson et al. (Mol. Microbiol. 36, 679-687 (2000)) reported that mutants deprived of Ycf1p displayed enhanced resistance to sodium selenite and suggested that Ycf1p mediated vacuolar transport of selenite under the form of a glutathione conjugate might deplete the reduced glutathione cytoplasmic pool (Pinson et al. (2000), Identification of genes affecting selenite toxicity and resistance in Saccharomyces cerevisiae, Mol. Microbiol. 36, 679-687). Therefore, we searched to know whether vacuolar Ycf1p transported selenodiglutathione (GS-Se-SG). For this purpose, we synthesized radioactive selenodiglutathione and prepared secretory vesicles harboring the Ycf1p protein. Results clearly showed that selenodiglutathione was a high-affinity substrate of Ycf1p. We also measured the GSH/GSSG ratio in selenite-stressed cells expressing Ycf1p and showed that it was 6-fold lower than in a strain deprived of Ycf1p. All these results indicate that transport of the selenodiglutathione adduct by Ycf1p decreases the reduced GSH cytoplasmic pool, a behavior which, in turn, increases the vulnerability of cells to the selenite-induced oxidative stress.
Uptake of selenite by the phosphate transporters (Lazard, M., Blanquet, S., Fisicaro, P., Labarraque, G. & Plateau, P., Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters. J. Biol. Chem. 285, 32029-32037 (2010) [Abstract] [Full text])
Although general toxicity of selenite has been extensively studied, little is known on the mechanism by which this compound crosses cellular membranes. At physiological pH, the selenite ion has a size and a charge similar to that of the phosphate ion. In addition, competition between selenite and phosphate uptake has been early observed in wheat roots. We therefore questioned whether selenite and phosphate could share a same pathway to enter S. cerevisiae cells.
S. cerevisiae cells possesses 5 carriers for phosphate uptake. Three of them are constitutively transcribed and take up phosphate with low affinity. The other two carriers are transcriptionally upregulated in response to phosphate starvation and display affinities for phosphate in the micromolar range. Using deletion mutants, we showed that both the high- and low-affinity phosphate transporters were involved in selenite uptake and that phosphate and selenite indeed compete for uptake by both these carrier systems. Inhibition by phosphate of the import of selenite has already been observed in plants, in the green alga Chlamydomonas reinhardtii and the yeast Candida albicans. Opportunistic transport of selenite by phosphate transporters seems therefore to be a general mechanism, at least in plants, algae and fungi.
Perspectives
Currently, our research interests are focused on the mechanisms of toxicity of organic selenometabolites, such as selenomethionine and selenocysteine, which are the main components in the dietary supplements administrated against cancer. Are these amino acids metabolically transformed to a common intermediary such as H2Se, or does their toxic action proceed through specific, as yet unidentified, pathways?