
Membrane and transport
Any machine (thermal engines, living organisms, etc.) basically consists in locally reducing entropy to the detriment of a larger whole. This involves dividing the space into partly permeable compartments. In many devices (cells and batteries, living cells ...), this role is provided by membranes whose porosity allows controlled transport of molecules from one compartment to another. We experimentally study this transmembrane transport from a fundamental point of view on synthetic membranes (made of polymer or alumina) or mimicking biological membranes (lipid bilayers).
Science often proceeds by analogy. Thus, a category of unidirectional molecular transports based on mechanisms similar to those of a ratchet has become a subject of study in itself in statistical physics (cf. P. Hänggi, F. Marchesoni. Artificial brownian motors: Controlling transport on the nanoscale. Rev. Mod. Phys., 81:387, 2009). These mechanisms are called Brownians if they work thanks to the thermal energy which is paradoxical because they tend to reduce entropy. The two examples below, taken in very different areas, fall into this category. From an electrical point of view, these are ionic current diodes that operate thanks to the thermal agitation.
Synthetic conical pores
The motion of ions through a membrane provided with channels (pores) results from forces which are generally exerted directly and individually on the ions, such as those originating from a transmembrane electric field. But theoretically, the balance of these forces also involves a term of a completely different nature. Indeed, when the transit is slow enough so that ions have time to explore transversely by Brownian motion the channel, longitudinal variations of its section produce a gradient of entropy from which a force emerges. Thus, when the channel has an asymmetrical shape (eg. Funnel-like), this effect can induce a non-return mechanism similar to a ratchet and pawl device or to the valve of a pump operating at a molecular level: ions go ahead in the pore but hardly come back.
To verify this idea, we designed and realized an experiment based on the following principle: this entropic force depends on the size of the ions; thus, when it is very different for anions and cations, conical pores can be expected to correct an alternating ionic current. This is precisely what we have observed by making membranes with chemical etching of heavy ions tracks in polymer films and by using a dilute solution of polyacid as electrolyte.
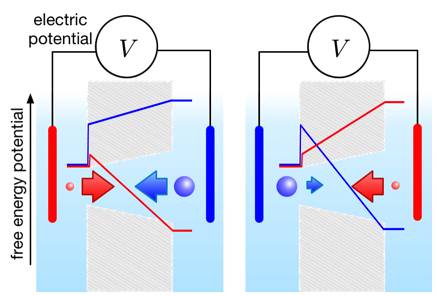
Schematic diagram of an entropy diode based on the geometric shape of the channel.
To know more:
The concept of entropic rectifier facing experiments, Sci. Rep., 6: 38966, 2016
Peptide pores of biological membranes
The interaction of biological cell membranes with molecules adsorbed on their surface and the way the incorporation of these molecules takes place are still poorly understood. We are interested in the antimicrobial peptides that are the basis of the innate immune system of multicellular organisms. Basically, these peptides make the membranes permeable by pore-forming. Their universal presence, their nonspecific and broad-spectrum action as well as their very elementary structure suggest a mode of action according to physical mechanisms that are also very general and universal.
We have addressed this problem by means of so-called "voltage-clamp" experiments (consisting of measurement of the transmembrane ionic current) and neutron reflectivity. In particular, we have shown that pore-opening displays a slow dynamics typical of the glass transition, which could be governed by surface fluctuations in peptide concentration as predicted by the RSA model (for Random Sequential Adsorption). More recently, we have been interested in the thermodynamics of pore opening as a function of transmembrane voltage and temperature. Our results allow us to propose a new physical mechanism where peptide-adsorption and electric field both tend to bend the membrane and in which the entropic cost of pore-opening results from an "excluded area" for the translation of lipids and explains how the necessary threshold voltage varies with temperature.
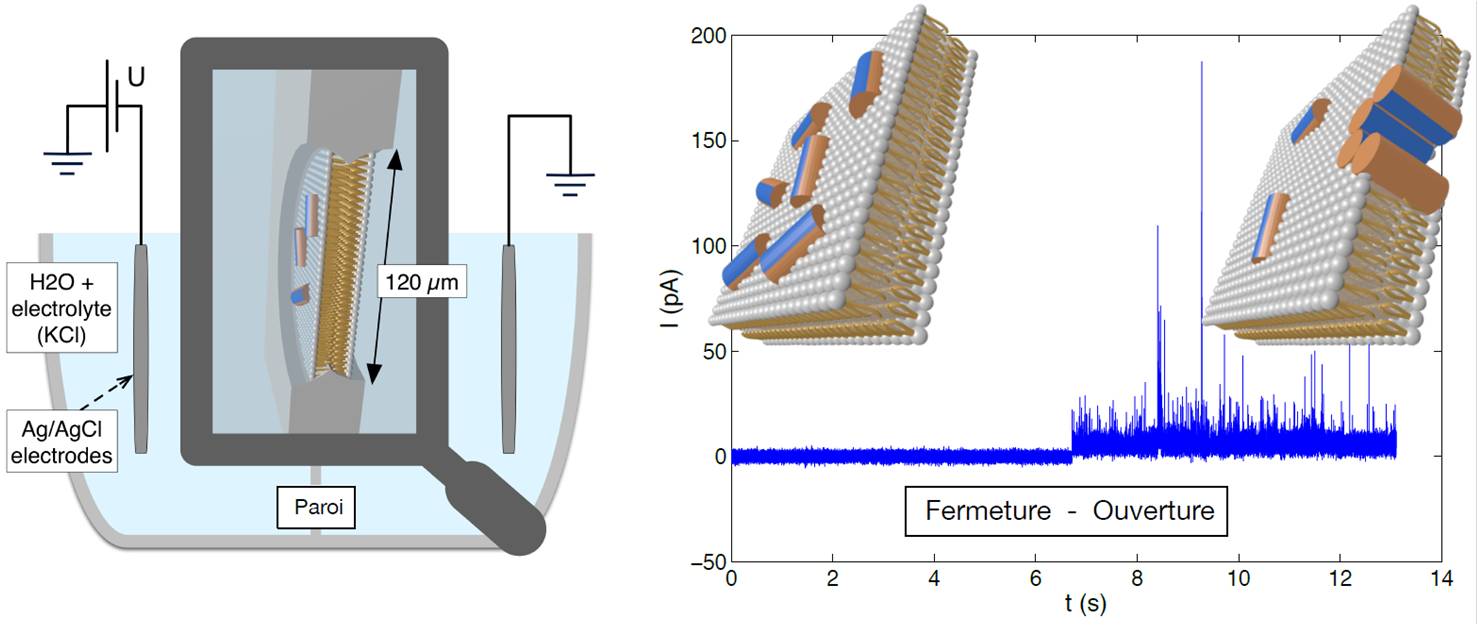
A self-supported membrane separates two electrolytes and allows us to detect pore-opening: to open/close corresponds conductor/insulator.
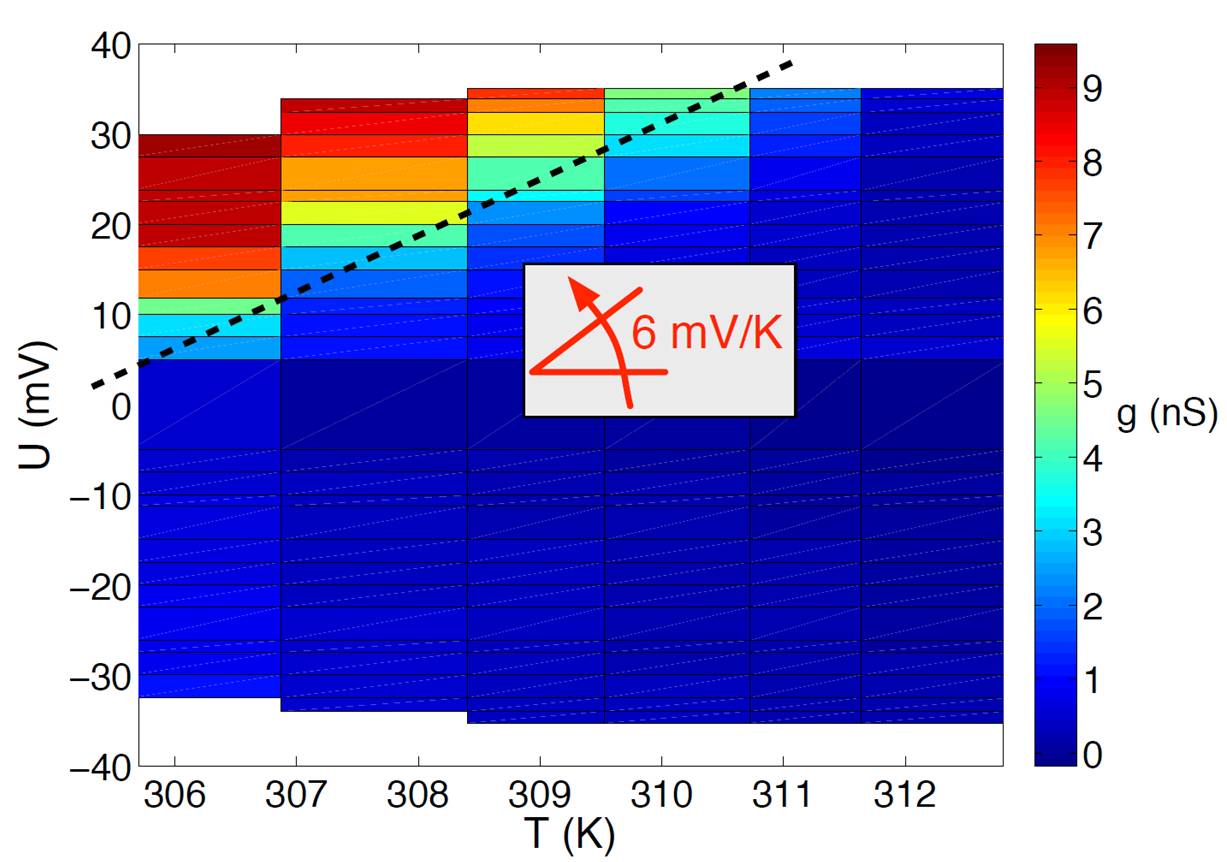
Conductance (g) of a lipid bilayer (DPPC) in the presence of the alamethicin peptide as a function of the potential U of peptide-compartment and of temperature T. The dotted line corresponds to the transition.
To know more:
Peptide pores in lipid bilayers : voltage facilitation pleads for a revised model, Phys. Rev. Lett., 111: 028102, 2013
Fluctuations of ionic current through lipid bilayer at the onset of peptide attacks and pore formation, Phys. Rev. Lett., 103: 180601, 2009